Data Zone
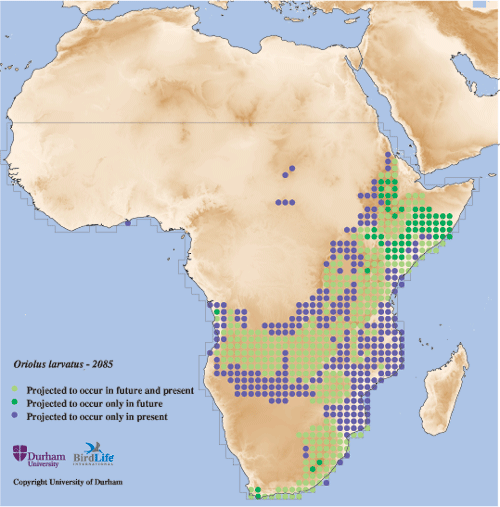 Climate change is expected to have a major impact on biodiversity, including on the distribution of species (see here to learn more). Through innovative collaborations using ‘climate envelope modelling’, BirdLife is researching where bird species are projected to occur in future. Here you can view maps of the projected distributions of bird species under climate change. At present, these maps are available only for bird species breeding in sub-Saharan Africa, and can be viewed by clicking on the ‘Climate Change Maps’ tab of each species factsheet.
Climate change is expected to have a major impact on biodiversity, including on the distribution of species (see here to learn more). Through innovative collaborations using ‘climate envelope modelling’, BirdLife is researching where bird species are projected to occur in future. Here you can view maps of the projected distributions of bird species under climate change. At present, these maps are available only for bird species breeding in sub-Saharan Africa, and can be viewed by clicking on the ‘Climate Change Maps’ tab of each species factsheet.
Alternatively, a full list of the assessed species can be viewed here.
Sample map for Mocking Cliff-chat Thamnolaea cinnamomeiventris
The ‘climate envelope’ of a species represents the association between its present-day distribution and current climatic variables. Future distributions are then estimated by projecting this relationship onto scenarios of climate change, making the assumption that current relationships between climate and distribution are retained (see e.g. Pearson & Dawson, 2003). As such, these maps represent the distribution of areas of potential climatic suitability for a species, rather than an explicit representation of each species realized distribution. In most cases however, there is likely to be a close relationship between the modeled distribution and the species ‘observed’ distribution.
The reliability of climate-envelope models has been extensively evaluated over the past two decades. They have been successfully used to model non-native/invasive species (e.g. Beerling, Huntley & Bailey, 1995; Menendez et al., 2006), recreate historical shifts in species distributions (e.g. Araujo et al. 2005, Nogues-Bravo 2009), and to detect consequent shifts in individual species’ population trends (e.g. Green et al., 2008; Gregory et al., 2009). However, these models are also subject to a range of assumptions and there is an acknowledged need for their continued development, improvement and testing (e.g. Araujo & Luoto 2007, Beale et al. 2008; Elith & Graham 2009, Guisan & Thuiller 2005, Hijmans & Graham 2006, Kearney & Porter 2009, Pearson et al. 2003, 2006). Any map depicting a species’ modelled distribution therefore needs to be interpreted carefully and judiciously. The maps on this website represent modelled projections of the distributional changes that could occur under climate change, not predictions of what will occur. As such they should be used as a tool to help scientists, policy-makers and other stakeholders identify and address potential adaptation needs, but within an adaptive management and decision framework (e.g. Hole et al. 2011) that includes other independent data sources on likely climate change impacts, and that explicitly recognizes the inherent uncertainties when addressing climate change.
Maps are currently available for 1608 species: the entire breeding avifauna of sub-Saharan Africa, minus 71 species recorded from fewer than five grid cells, for which modelling was impractical. The maps depict modelled present-day and future species ranges approximating to four discrete time periods: present-day (based on the mean climate between 1970–2000), 2025 (a mean of climate projections for the period 2010–2039), 2055 (mean for 2040–2069) and 2085 (mean for 2070–2099). The models are based on comprehensive ‘observed’ distributional data for all terrestrial bird species breeding in sub-Saharan Africa, provided by the Zoological Museum of the University of Copenhagen (see here). The continental landmass south of latitude 20°N is divided into 1963 1° x 1° latitude/longitude cells, of approximately 111 km x 111 km at the Equator. The modelled distribution of each species is then indicated by its modelled presence or absence in these cells. Mean monthly temperature and precipitation data for sub-Saharan Africa were obtained from a global, freely available dataset (www.worldclim.org), while projections of future climate change were derived from three General Circulation Models (GCMs) included in the IPCC’s Third Assessment Report (all GCM data are available at www.ipcc-data.org). For further details, see Hole et al. (2009). Note that the time-scale for these projections is likely to prove conservative if the current CO2 emissions trajectory continues. Emissions growth rate since 2000 has been at the upper end (and has possibly even exceeded) the most fossil-fuel intensive of the IPCC emissions scenarios (Raupach et al. 2007), with a projected temperature increase in the range of 2.4–6.4°C, in comparison to 1.4–3.8°C for the scenario used here. Should this trend continue, then it is likely that these projections for avian turnover and persistence for 2085 will be manifest on a shorter time-scale, perhaps as early as 2055.
References
Araujo, M.B. & Luoto, M. (2007) The importance of biotic interactions for modelling species distributions under climate change. Global Ecology and Biogeography, 16, 743-753.
Araujo, M.B., Pearson, R.G., Thuiller, W., & Erhard, M. (2005) Validation of species-climate impact models under climate change. Global Change Biology, 11, 1504-1513.
Beale, C.M., Lennon, J.J., & Gimona, A. (2008) Opening the climate envelope reveals no macroscale associations with climate in European birds. Proceedings of the National Academy of Sciences of the United States of America, 105, 14908-14912.
Beerling, D.J., Huntley, B., & Bailey, J.P. (1995) Climate and the Distribution of Fallopia-Japonica - Use of an Introduced Species to Test the Predictive Capacity of Response Surfaces. Journal of Vegetation Science, 6, 269-282.
Elith, J. & Graham, C.H. (2009) Do they? How do they? WHY do they differ? On finding reasons for differing performances of species distribution models. Ecography, 32, 66-77.
Green, R.E., Collingham, Y.C., Willis, S.G., Gregory, R.D., Smith, K.W., & Huntley, B. (2008) Performance of climate envelope models in retrodicting recent changes in bird population size from observed climatic change. Biology Letters, 4, 599-602.
Gregory, R.D., Willis, S.G., Jiguet, F., Vorisek, P., Klvanova, A., van Strien, A., Huntley, B., Collingham, Y.C., Couvet, D., & Green, R.E. (2009) An Indicator of the Impact of Climatic Change on European Bird Populations. Plos One, 4.
Guisan, A. & Thuiller, W. (2005) Predicting species distribution: offering more than simple habitat models. Ecology Letters, 8, 993-1009.
Hijmans, R.J. & Graham, C.H. (2006) The ability of climate envelope models to predict the effect of climate change on species distributions. Global Change Biology, 12, 2272-2281.
Hole, D. G., Huntley, B., Pain, D. J., Fishpool, L. D. C., Butchart, S. H. M., Collingham, Y. C., Rahbek, C. and Willis, S. G. (2009) Projected impacts of climate change on a continental-scale protected area network. Ecol. Lett. 12: 420-431.
Hole, D. G., Huntley, B., Collingham, Y. C., Fishpool, L. D. C., Pain, D. J., Butchart, S. H. M. and Willis, S. G. (2011) Towards a management framework for protected area networks in the face of climate change. Conserv. Biol. 25: 305–315.
Kearney, M. & Porter, W. (2009) Mechanistic niche modelling: combining physiological and spatial data to predict species' ranges. Ecology Letters, 12, 334-350.
Menendez, R., Megias, A.G., Hill, J.K., Braschler, B., Willis, S.G., Collingham, Y., Fox, R., Roy, D.B., & Thomas, C.D. (2006) Species richness changes lag behind climate change. Proceedings of the Royal Society B-Biological Sciences, 273, 1465-1470.
Nogues-Bravo, D. (2009) Predicting the past distribution of species climatic niches. Global Ecology and Biogeography, 18, 521-531.
Pearson, R.G. & Dawson, T.P. (2003) Predicting the impacts of climate change on the distribution of species: are bioclimate envelope models useful? Global Ecology and Biogeography, 12, 361-371.
Pearson, R.G., Thuiller, W., Araujo, M.B., Martinez-Meyer, E., Brotons, L., McClean, C., Miles, L., Segurado, P., Dawson, T.P., & Lees, D.C. (2006) Model-based uncertainty in species range prediction. Journal of Biogeography, 33, 1704-1711.
Raupach, M.R., Marland, G., Ciais, P., Le Quere, C., Canadell, J.G., Klepper, G., & Field, C.B. (2007) Global and regional drivers of accelerating CO2 emissions. Proceedings of the National Academy of Sciences of the United States of America, 104, 10288-10293.